 北野成昭(キタノナルアキ) 作『獣医志Wiki』はクリエイティブ・コモンズ 表示 - 非営利 - 継承 4.0 国際 ライセンスで提供されています。
北野成昭(キタノナルアキ) 作『獣医志Wiki』はクリエイティブ・コモンズ 表示 - 非営利 - 継承 4.0 国際 ライセンスで提供されています。
細菌学総論:細菌の一般性状
目次
染色法
1、普通染色法
- 1単染色→メチレンブルーなど
- 2.グラム染色→陰性菌と陽性菌のペプチドグリカンの構造の違いを利用して染め分ける方法
- 塗沫、乾燥、固定後の標本をクリスタルバイオレット(青紫)で染色しルゴール液で触染、エタノールで脱色後にサフラニン液(赤)もしくはPfeiffer液(赤)で後染色する。
- グラム陽性菌はペプチドグリカン層が厚く資質に乏しいためエタノールで処理しても細胞壁の透過性は低いままなので、青紫に染まったままである。
- グラム陰性菌は陽性菌とは逆で細胞壁に脂質が多いためエタノール処理で脂質が溶解して細胞壁の透過性高まり、クリスタルバイオレット・ルゴール複合体が抽出されて脱色される。よってサフラニンに染色される。
2、特殊染色
検鏡法
- 光学顕微鏡
- 電子顕微鏡
細菌の微細構造
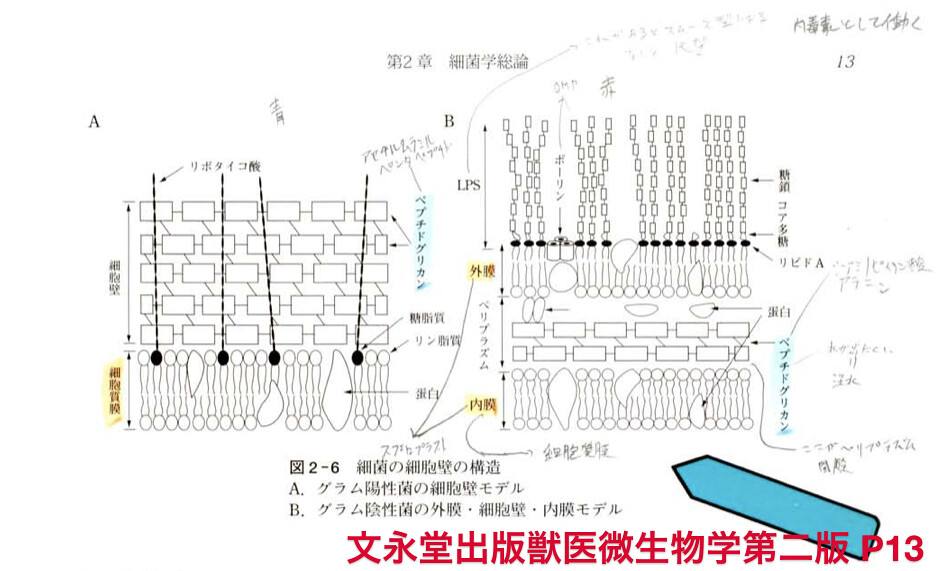
細胞壁
1.グラム陽性菌:グラム陽性菌の細胞膜の外側は強固な厚い細胞壁で覆われ,細胞壁はペプチドグリカンタンパク,タイコ酸(リポタイコ酸も、ペプチドグリカンの構造を補強している),多糖から構成されている. プロトプラスト(細胞壁ない状態)
2.グラム陰性菌:グラム陰性菌の細胞膜の外側にはペプチドグリカン層があり,その外側を外膜が覆っている複雑な細胞膜構造をもっている.スフェロプラスト(細胞壁ない)
- LPSの生物活性
- LPSはグラム陰性菌に特有のもので,動物に対して毒素活性をもち,内毒素(エンドトキシン)と呼ばれています.通常,細胞壁に強固に結合していますが,細胞の溶解が起こったときにのみ遊離し,生体に様々な反応を引き起こします.
- 内毒素はタンパク毒素である外毒素と異なり,熱,乾燥,消毒剤に対して強い抵抗性を持ちます.活性の作用はリピドA部分のものがほとんどです.しかしリピドAそのものは水に不溶ですのでそれだけでは活性を示しません.多糖部分の大きな親水性により可溶化して,水溶液中ではミセルを形成して運ばれれ活性を示します.
- LPSは補体を第2経路において活性化させ,マクロファージや他の細胞からサイトカインや他の免疫調整物質を放出します.これらの生理活性物質は宿主の生体防御機構の正常な作用とそていは細菌を除去する方向に働きます.しかし,これらのサイトカインなどが過剰になると宿主に毒性をしめし,ショックなど死を招くことがあります. LPSにようり細胞が刺激されるとサイトカイン
外膜
外膜はグラム陰性薗にのみ見られ,リン脂質,タンパク,リボタンパク,リボ多糖で構成される強埼な構造で,ホスホリバーゼA,フアージレセプター,コリシンなどが存在する.外膜は内膜(細胞質膜)とは異なり,含有する酵素の種頼も内膜ほど多くはない.外膜タンバタの種類は内膜に比べて桁違いに少ない.大腸菌の場合,主要な構成タンパクとしては分子量40,000前後の主要外膜タンパク(大腸菌でOmpA,OmpC,OmpFなど)や特殊な修飾をN末端にもつ主要リポタンバタなどが存在する.それぞれのタンパク量は非常に多く,いずれも菌体に存在する最も多いタンパクである.最近,環境の変化に対して,外膜タンバタの発現がダイナミックに変化することが知られるようになり,環境の変化に機敏に反応し,その構成を変え,外界から細胞を守る重要な役割を果たしていると考えられる.多くの細菌に存在するポリンタンパクや大腸菌の主要外膜タンパクのOmpC,OmpFは三量体を形成し,外膜を貰通している.これらは低分子親水性物質に対する透過孔を形成し,栄養低分子,抗生物質の外膜透過に関与している.また,病原性,築剤の透過性や薬剤耐性を担う機構も外膜に存在する.これらの主要外膜タンバタの発現は環境の変化に対してきわめて敏感に反応し,合成制御されている.
- 外膜は酵素などのタンパクの種類が少なく,またペプチドグリカンと強固に結合し ているところから機能的に細胞壁と同じ意味に扱われることが多い.しかし,現在で は多くの機能タンバタの存在が知られるようになり,また膜構造からも単位膜である.
細胞質膜
- 選択的膜透過性と物質輸送
- 電子伝達系と酸化的リン酸化
- 酵素や代謝産物の分泌
- 細胞壁合成、脂質合成
細胞質膜はリン脂質とタンパクで構成され,呼吸,酸化的リン酸化をつかさどるエ ネルギー合成機構,栄養物などを取り込む能動輸送(activetransport)に関する機構, 外界の刺激を受け取るレセプターとその信号を細胞の中に伝える機構などが存在し, 基本的に動物細胞の細胞膜に相当する.細胞質膜にはプロトンボンプが存在し,これ によってつくられた陽イオン勾配と膜電位に貯蔵されたエネルギーによってアミノ酸 や糖のような溶質を細胞内に能動輸送する.膜に存在するキヤリヤータンバタが陽イ オンと溶質に結合して,両者を同時に輸送する.
- 細菌ではシトクロムやその他の呼吸連鎖の酵素は細胞質厳に局在し,真核細胞のミ トコンドリアの内膜に非常に類似している.このことがミトコンドリアが共生細菌か ら進化したものであると考える説の根拠の一つとなっている.
また,細胞質膜からは多くの水解酵素が分泌され,栄養の取込みなどに役立ってい る.グラム陰性菌では外膜と細胞膜(内膜)との間にある間隙(ペリプラズム)にこれらの酵素が曹積され効率的に働いている.また,細胞質膜にはこれらの分泌タンバタを 分泌する機構がある.分泌タンバタを,そのN末端の特異なペプチド(シグナルペプチ ド)を介して細胞質膜の外に押し出し,分泌後,シグナルペプチドを切断するシグナル ペプチダーゼも細胞質膜に存在する.その他,細胞質膜には鞭毛を回転させるモータ ー機構や,外界の環境を察知するセンサータンパク,その情報をタンバタ合成系に伝 達するタンバタがあり,環境変化に適合させる各種のタンパクの合成が開始される. 外膜タンパクOmpC,OmpFも浸透圧の変化によって発現を制御されている代表的な タンパクである.
プチドグリカン
- 細菌は動物細胞とは違って、細胞の外層は堅い細胞壁で被われています。細菌の形が一定に保たれ、外圧や浸透圧の変化などの外界の環境の変化にも対応しいるのもこの細胞壁のおかげです。この細胞壁に一定の堅さを与えているものがペプチドグリカンで、格子構造で菌全体を袋状に覆っています。
- ペプチドグリカンは多糖グルカン鎖を縦糸に、ペプチドの側鎖を横糸にして、格子状に織り上げた布のような三次元的な網目構造をしています。
- 縦糸である多糖グルカゴンはN アセチルグルコスアミン(GluNAc)とN アセチルムラミン酸 (NurNAc)がβ 1 4結合したものです。
- 横糸であるペプチドはアミノ酸が4つ結合したもの(テトラペプチド)です。
グルカン鎖が菌体の長軸にたいして垂直にならび、取り囲むように走行して、隣のグルカン鎖との間にペプチド結合で結合しています。すなわち,グリカン鎖のMurNAcに結合したテトラペプチドは,別のグリカン鎖結合しているテトラペプチドに直接結合するか(大腸菌の場合),あるいは他のアミノ酸鎖(黄色ブドウ球菌の場合は5分子のグリシン)を介して結合しています.この結合は立体的に積み重ねられて層を形成しているのです.
- グラム陽性菌では外膜の40~70%がペプチドグリカンですが、グラム陰性菌での割合は数%にしかすぎません。グラム陰性菌では外膜の内側に裏打ちされており、外膜とともに細胞壁を形成してます。
- 涙や卵白などに含まれるリゾチームはペプチドグリカンを破壊する働きのある酵素で、病原菌から生体を守る働きをしています。しかしグラム陰性菌ではペプチドグリカンがリゾチームによって破壊され、細胞壁は脆弱になりますが、胞壁の主成分は外膜ですので細菌の形は保たれます。
また、ペプチドグリカンは細胞壁に作用するペニシリン系、セフェム系の抗生物質の作用部分として重要す。(ペニシリンは)ペンタペプチド末端のD アラニール D アラニンの構造に類似しており、ペプチドグリカン合成酵素に選択的に結合し、架橋構築を阻止し、その結果菌が破壊されるのです。
- 核
- 核膜なし
- メソソーム:リボソーム(30Sと50Sのサブユニットが会合して70Sのリボソームを形作る)
- 異染小体:高エネルギー源、リン酸源の貯蔵庫と考えられている。
- プラスミド:染色体DNAとは別個に自己複製する環状二本鎖DNA宿主菌の発育・増殖に不可欠なものではないが重要な表現形がここにコードされている。
莢膜
- 菌の種類によっては細胞壁の外側に透明な層が取り囲んでいる部分。
- 境界が不明瞭なら粘液層、明瞭なら莢膜
芽胞
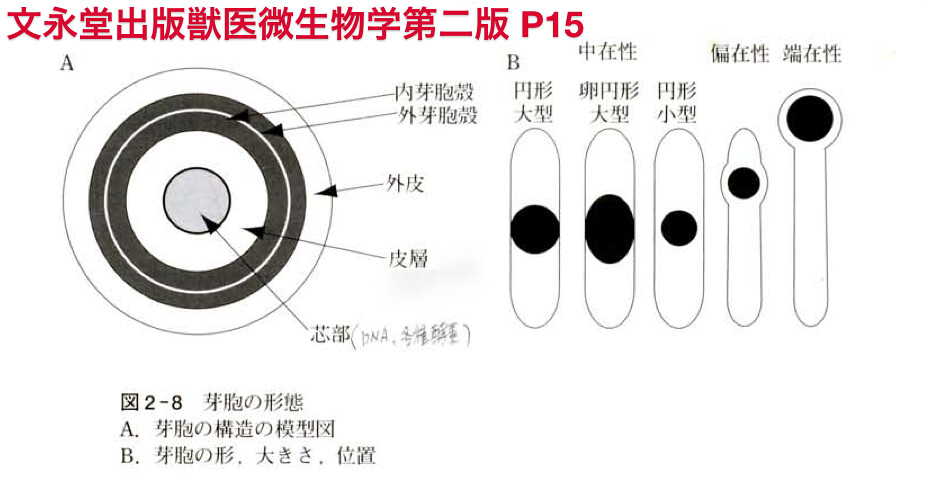
- 芽胞を形成するBacillus cereus やClostridium
- 硬い皮膜で桿菌をおおい、生物活動を休止させる。
- 細菌細胞は周囲の環境がその細菌の生育に不利な状況になると死滅していくのが一般的です.しかし,ある種の細菌は乾燥,高温などの環境条件が悪くなると芽胞とよばれる耐久器官を作り生き延びていきます. この芽胞を形成する菌で好気性の細菌をBacillus属,嫌気性の細菌をClostridium属と呼んでいます.両者ともグラム陽性の比較的大きい桿菌です. 芽胞形成菌であっても栄養バランスがとれていて分裂増殖しているとき(栄養型)には,芽胞を形成しないときがおおく,バランスがくずれたときに菌体内に芽胞を生じます(スポランギウム) 芽胞では代謝が行われていない休止状態で細菌の増殖などの生物活性がほとんど見られません. (芽胞は細菌胞子とか内生胞子とかよばれることもあります)
耐性
- 物理的,化学的刺激に対して強い抵抗性を持っています. 水分の少ない濃厚な原形質と核をあつい殻で覆っており,乾燥熱,消毒剤のような化学薬品処理,紫外線,放射線照射に対して強い抵抗性を示します. 120℃15分のオートクレーブであらゆる芽胞は完全に死滅しますが,100℃の煮沸にも耐えられます. 芽胞はエタノール消毒には耐えるために芽胞の汚染が考えられる場合の消毒にはヨードや塩素系での消毒が必要となります. 遊離した芽胞は特に耐性で,数10年にわたり生存することができます.
- 芽胞は菌種によって菌体内にできる位置が異なっています.形は破傷風菌の芽胞は球形ですが枯草菌の芽胞は楕円型です. 芽胞は屈折性の強い物質であり,位相差顕微鏡などでみると,芽胞だけが強く光って見えます. 菌1個あたり出来る芽胞の数は通常1個ですが,まれに2個のときもあります. 芽胞は光を通さず,染色もされにくい性質をもち,グラム染色や単染色では抜け像として観察されます. 石炭酸フクシンを使った芽胞染色法によって観察できます.
成分
また,芽胞ではその水分の60 70%は結合水の形になっており自由水をもたない脱水状態になっています. 芽胞の5 12%を占めるジピコリン酸は,大量のCaを持っています.Ca,Cuなどを多く含有しています. 酵素はあるがATPはほとんど持たず,NADやFADは不活性な状態にあるためです. 芽胞は温度,湿度が適当になると増殖型(栄養型)細菌となります. 細胞分化のモデルともなっています.
過程(内容)
- 芽胞形成 炭素,窒素源が欠乏し,栄養条件がととのってない条件下で,対数増殖期が終了した時期に始まります.また,芽胞形成には,栄養細胞機能に関連する遺伝子の不活性化にともない,芽胞形成遺伝子の活性化がおこります.それにともないこの時期に特有の遺伝子産物として新たな酵素や代謝物が作られ,芯部(核,細胞質)ができ,皮層ができると同時に,染色体を凝集してリボソームとタンパクの一部を濃縮し硬い被膜で覆い包み,呼吸などの代謝はほとんど行わず生物活性をほぼ完全に休止させます. 培養が古くなると母細胞が崩壊し去り,残された芽胞は遊離し完成された芽胞となります. 新しい構造が作られていきます.生物活性は休止します.
発芽 酸化などの要因によって芽胞殻に損傷が加えられた場合とか,グルコースやアラニンのような発芽剤と接触すると発芽の活性化がおこります. 芽胞殻変性後,芽胞の融解,水分の取り入れなどの芽胞変性をへて細胞質から新たな栄養細胞が出現します.
細菌の栄養
栄養型サイクル 栄養,環境が好転すると再び分裂増殖をはじめます.
- 自力(自家)栄養菌→無機物を栄養源として増殖する菌
- 光合成自力栄養菌
- 化学合成自力栄養菌
- 従属栄養細菌→エネルギー源として何らかの有機物を必要とする。獣医学領域で重要な病原菌はこれに該当する。
- 炭素源:co2 HCO3は自家栄養菌。従属は炭水化物、アルコール類、アミノ酸類、ペプチド、有機酸を炭素源とする
- 窒素源:無機物としてはNH4,NO3,NO2、有機物としては蛋白、ペプチド、アミノ酸が利用される。
- 無機塩類:Pは核酸やATP、Sはアミノ酸合成に不可欠。Na,K,Mg,Ca.Fe.Cu.Zn.Co.Clは水素イオン濃度や浸透圧のために必要である。
- 発育因子:補酵素もしくはその前駆体となるビタミンB群など。